
Jedes Jahr im Herbst ist es soweit: Die Jahrestagung der Wirbeltierpaläontologen (ausgerichtet von der Society of Vertebrate Paleontology) findet statt. Normalerweise lese ich allenfalls darüber, aber dieses Jahr war die Tagung in Berlin – und das konnte ich mir natürlich nicht entgehen lassen. Und so habe ich die letzten 4 Tage (mit einer kurzen Rückkehr nach Braunschweig, weil ich noch eine Vorlesung zu halten hatte) in Berlin verbracht und mir dort Vorträge angehört, Poster angeschaut (allerdings waren die Poster-Sessions sehr eng, so dass man viele Poster ohne massiven Ellbogeneinsatz nicht richtig angucken konnte) und mit einigen Leuten geredet.
Wenn ich richtig gezählt habe, habe ich 67 Vorträge gehört (kein Wunder, dass mir abends der Kopf schwirrte) – über die Abschätzung der Masse von Vögeln, über die Knochenhistologie von Temnospondylen, über die Biomechanik von Sauropodenwirbeln, die Evolution von Schildkröten, und und und.
Nein, über alle werde ich hier wohl eher nicht schreiben, aber hier ein kleiner Überblick über die, die mir am meisten im Gedächtnis geblieben sind. (Wenn es euch interessiert, könnt ihr euch das “Book of Abstracts” herunterladen und lesen, was es sonst noch so gab.)
Los ging das Ganze für mich am Mittwoch Vormittag gegen 11:00Uhr im Archaeopteryx-Symposium, in dem es aber nicht nur um den Urvogel, sondern generell um die Evolution der Vögel ging. Der erste Vortrag, den ich vollständig gehört habe, beschäftigte sich mit der Frage, ob wir wirklich die Farben von fossilen Federn rekonstruieren können – dazu gab es ja in den letzten Jahren einige Arbeiten, die fossile Melanosomen angeschaut haben (Melanosomen werden von Zellen produziert und sind kleine, etwa kugelförmige Gebilde, die den Farbstoff Melanin enthalten). Dazu wurden Vogelfedern mehrere Jahre lang unter verschiedenen Bedingungen (bei 60°, bei 350°, in Sand eingebuddelt) gelagert und dann geschaut, was nach dieser Zeit von den Melanosomen und den Keratinfasern drum herum noch übrig ist. Ihr Fazit (leicht vereinfacht): Melanosomen werden schneller zerstört als Keratinfasern – wenn man also Teile findet, die wie Melanosomen aussehen, aber keine Keratinfasern mehr finden kann, dann sind die Melanosomen vermutlich gar keine, sondern etwas anderes (beispielsweise Bakterien). Das Ergebnis dürfte einigen Leuten nicht so gut gefallen – mal sehen, wie diese Debatte weitergeht.
Danach ging es um Embryologie – genauer um die Herkunft des Vogelschnabels. Der Vogelschnabel sitzt auf einem Knochen, der evolutionär durch das Verschmelzen der beiden vorderen Kieferknochen (Praemaxillae) entstanden ist. Dann wurde an Embryonen untersucht, welche Steuergene bei Vögeln (anders als bei anderen Tieren) in diesem Bereich aktiv sind und schließlich wurden diese Gene dann gezielt ausgeschaltet – mit dem Ergebnis, dass die entstehenden Embryonen wieder die ursprünglichen Anlagen der Kieferknochen zeigen. Der Schritt vom Saurierkiefer zum Vogelschnabel ist also zumindest genetisch vielleicht gar nicht so kompliziert, wie man gedacht hätte.
Anschließend zwei Vorträge über die Entwicklung der Vögel im Allgemeinen – der Vortrag von Benton (einem der ganz großen Namen in der Wirbeltierpaläontologie, dessen Buch ein Standardwerk ist, dass in keinen Haushalt fehlen sollte) war eher allgemeiner Natur (und war zwar nett vorgetragen, ist mir aber ehrlich gesagt wenig in Erinnerung geblieben); der anschließende Vortrag beschäftigte sich dann mit der Frage, wie schnell sich die Vögel an unterschiedliche ökologische Nischen angepasst haben. Es zeigt sich, dass es schon im Eozän (so vor rund 50 Millionen Jahren) eine große Vielfalt an unterschiedlichen Vögeln gab. Das oft gezeigte Bild, wonach die Vielfalt der Arten im Laufe der Evolution bis zum heutigen Tag (von einigen Aussterbe-Ereignissen abgesehen) immer weiter zunahm, ist also vermutlich nicht ganz zutreffend.
Und als letztes ging es dann vor der Mittagspause noch darum, in wie weit es möglich ist, die Massen von Vögel (speziell Wasservögeln) an Hand ihrer Knochen vorherzusagen – es zeigt sich, dass die Länge der Arm- und Beinknochen kein gutes Maß ist (wenig überraschend, da es ja sehr lang- und kurzbeinige Vögel gibt); der Umfang der Knochen (insbesondere des Unterschenkels) erlaubt dagegen eine einigermaßen guten Schätzung. Ich finde das ein bisschen überraschend, weil ja längere Knochen auch größere Biegemomente bedeuten und deswegen auch der Querschnitt der Knochen größer sein sollte – aber vielleicht kompensieren das die langbeinigen Vögel ja dadurch, dass sie ihre Beine gerader halten. (Der Vortrag selbst beschäftigte sich nur mit lebenden Vögeln – trotzdem gehört er natürlich auf eine SVP-Tagung, weil man ja irgendwie überlegen muss, wie man Massen ausgestorbener Tiere abschätzen kann.)
Nach der Mittagspause kam die Theropoden-Session (o.k., parallel gab es noch andere über Säugetiere und Amphibien…). Gespannt war ich vor allem auf den Vortrag zum Spinosaurus – der war aber in gewisser Weise enttäuschend. Nizar Ibrahim hat zwar einen Super-Vortrag gehalten, der an Professionalität kaum zu überbieten war (und von daher war es natürlich cool), aber es war ein Vortrag, wie er ihn auch für ein allgemeines Publikum hätte halten können und der dieselbe geschichte erzählte, die man vor ein paar Wochen bei Terra X sehen konnte – ich hätte mir ja etwas mehr Details dazu gewünscht, wie genau man denn nun die unterschiedlichen Fossilien und die Zeichnungen von Stromer zu einem Gesamtbild zusammengesetzt hat und welche Methoden da eingesetzt wurden.
Danach ging es um Bruchmechanik – und vermutlich gab es außer mir nicht viele im Publikum, die etwas mit Spannungsintensitätsfaktoren und J-Integralen anfangen konnten… Das eigentliche Problem habe ich vor langer Zeit schon mal erklärt: Es geht um die Kerben an Raubsaurierzähnen und die kleinen Ausrundungen (Ampullae) an den Enden der Kerben. In der neuen Untersuchung wurden zum einen Simulationsrechnungen gemacht (das zweidimensionale Modell des Dinozahns fand ich aber nicht überzeugend – da der Querschnitt so eines Zahns ja nicht gerade flach ist, ist eine 2D-Ansicht vermutlich quantitativ ziemlich weit von der Realität entfernt. Zum anderen wurde gezeigt, dass das Material am Grund der Kerben typischerweise andere Eigenschaften hat und deswegen zusätzlich als Riss-Stopper dienen kann.
Zwei Vorträge befassten sich mit dem Nistverhalten von Dinos. Zum einen ging es um Oviraptorosaurier – die zweibeinigen Raubsaurier, von denen man früher dachte, dass sie Eier gefressen haben, weil man sie oft zusammen mit Nestern gefunden hat – während man jetzt weiß, dass es ihre eigenen Nester waren, die von ihnen bebrütet wurden. (Ein bisschen was über diese Funde findet ihr in einem meiner persönlichen Lieblingsartikel auf diesem Blog). Oviraptorosaurier gab es in sehr verschiedenen Größen – die meisten waren eher klein, aber Gigantoraptor erreichte eine Länge von etwa 8 Metern. Haben alle diese Dinos ihre Eier ausgebrütet oder haben die großen vielleicht doch eher Nester gebaut, wie es Krokodile tun, in denen die Eier dann durch zerfallendes Pflanzematerial gewärmt werden? Dazu hat man sich zunächst angeschaut, wie porös die Schalen waren: In bedeckten Nestern verlieren die Eier weniger Wasser und haben deswegen mehr oder größere Poren (ich hoffe, ich erinnere mich richtig an den Zusammenhang). Schaut man sich die Porosität der Schalen an, dann zeigt sich, dass alle Eier, die vermutlich von Oviraptorosauriern stammten, ausgebrütet wurden. Stellt sich natürlich die nächste Frage, wie ein mehrere Tonnen scherer Saurier wie Gigantoraptor das anstellt, ohne die Eier beim Draufsetzen in Rührei zu verwandeln – aber es zeigt sich, das bei größeren Nestern in der Mitte des Nestes immer ein bisschen Platz frei bleibt, so dass die Eier eher ringförmig angeordnet sind, so dass der größte Teil des Gewichts auf dem Boden lag. (Bei kleinen Sauriern war das anders, weil die Eier im Verhältnis zur Körpergröße wesentlich größer waren, so dass die Tiere nicht groß genug waren, um in der Mitte eines Rings zu sitzen und trotzdem noch alle Eier warmzuhalten.
Der andere Vortrag zum Nestbau (mit dem schönen Titel “lay-brood-repeat” und angepasstem Kinoposter zum Abschluss) beschäftigte sich mit den Nestern von Troodontiden, eher kleinen Raubsauriern vom Ende der Kreidezeit. In deren Nestern findet man oft zwei Lagen Eier übereinander, von denen die unteren nur in Bruchstücken vorhanden sind. Das lässt darauf schließen, dass die Troodontiden ihre Eier wiederholt in dieselben Nester gelegt haben (und es wurde auch ein bisschen überlegt, wie genau das abgelaufen sein könnte).
Danach gab es noch die Postersession und dann musste ich – wegen meiner Vorlesung am Donnerstag – zurück nach Braunschweig. (Dass die Lokführer zwischendurch gestreikt haben, machte die ganze Reiserei nicht unbedingt einfacher – aber so habe ich mal das deutsche Fernbus-System kennengelernt und dank hinreichend frühen Buchens auch einiges an Geld gespart.) Die Vorträge am Donnerstag habe ich dann leider auch komplett verpasst (was sehr schade war, weil am Donnerstag die Biomechanik-Vorträge lagen), aber ich kam gerade noch rechtzeitig zur Postersession. Das war nicht ganz unwichtig, denn diesmal (anders als vor drei Jahren in Obernkirchen) war ich nicht nur einfach Zuhörer, sondern hatte eigene Ergebnisse zu zeigen. Aber darüber schreibe ich ein andermal – die Veröffentlichung zum Thema erscheint ohnehin demnächst und dann erzähle ich euch, was ich da genau gemacht habe.
Freitag morgen gab es mal keine Session über Dinos, und so habe ich ein bisschen hier und da hineingehört – erst eine halbe Vortragssitzung über Temnospondylen (Verwandte der heutigen Amphibien), dann über Säugetiere. Bei den Temnospondylen ging es in mehreren Vorträgen um die Struktur ihrer Knochen und was man daraus schließen kann – das Thema hatten wir hier ja schon letztes Jahr. Außerdem wurden einige neue Fossilien vorgestellt (dazu schreibe ich nicht so viel, weil das ohne Bilder wenig ergiebig ist (Fotografieren war explizit untersagt – sonst könnte jemand Bilder im Vortrag machen und dann ein neues Fossil als erster veröffentlichen und benennen)). Außerdem gab es noch eine Analyse der Verbreitung unterschiedlicher Arten, in der geschaut wurde, in wie weit Temnospondylen zu welchen Zeiten endemisch waren (also an unterschiedlichen Orten mit nur dort zu findenden Arten vertreten waren) oder nicht und wie sich das mit anderen Tiergruppen vergleicht.
Anschließend habe ich dann bei den Säugetieren reingeschnuppert. Der erste Vortrag war in meinen Augen ein wenig seltsam – es wurden alle bekannten Großkatzen-Fossilien (und außerdem die Borhyaeniden) aus Europa räumlich und zeitlich eingeteilt und dann wurde geschaut, in wie weit sich über die Zeit Muster ergaben, bei denen Arten oder Gruppen durch andere ersetzt wurden. Statistisch schien mir das etwas fragwürdig – zum einen wurden für all die vielen untersuchten Muster immer nur p-Werte der Signifikanz angegeben (was mich hieran erinnerte), zum anderen frage ich mich, ob man bei etwa 300 Fossilien, die man in knapp 20 Zeiten aufteilt, die Fundlage wirklich gut genug ist, um daraus klare Schlüsse zu ziehen. Vielleicht hätte man mal spaßeshalber 10% der Fossilien zufällig aus der Stickprobe herausnehmen sollen und schauen sollen, ob sich dann etwas ändert. So schien mir das Ganze ein wenig als Zahlenspielerei. Solche eher statistischen Untersuchungen gab es einige in dieser Sitzung; mit den meisten davon konnte ich nicht so viel anfangen.
Faszinierend war aber der Vortrag über Tibet als “Eiszeit-Trainingslager”. Das tibetanische Hochplateau liegt zwar ziemlich weit südlich, ist aber trotzdem verdammt kalt und wird deswegen manchmal als dritter Pol der Erde bezeichnet. Tiere die dort leben, sind entsprechend an große Kälte angepasst – und das war auch vor der letzten Eiszeit so. Zumindest vom berühmten eiszeitlichen Wollnashorn weiß man inzwischen, dass seine Vorfahren in Tibet lebten und dort ihre Anpassungen an die Kälte erwarben. In dem Vortrag ging es darum, zu untersuchen, ob das auch für andere Arten gelten könnte. Hierfür gibt es inzwischen einige Hinweise – auch arktische Füchse hatten vermutlich ihre Vorfahren in Tibet, ebenso die eiszeitlichen Schneeleoparden und Hyänen. Die Idee von Tibet als Trainingslager für große Kälte könnte also auf einige Eiszeit-Tierarten zutreffen. (Und wirft natürlich die Frage auf, wie die Eiszeit-Fauna ausgesehen hätte, wenn es Tibet nicht gegeben hätte…)
Nachmittags ging es dann weiter mit urzeitlichen Reptilien – es gab ja nicht nur Dinos, sondern auch viele andere seltsame Viecher, insbesondere in der Zeit vor den Dinos. Eine gute Übersicht über die Verwandtschaftsverhältnisse findet ihr bei Wikipedia – hier ein Screenshot der Verwandtschaftsverhältnisse (aber Achtung: das Kladogramm ist nicht ganz aktuell und die Position der Schildkröten (Testudinae) ist höchstwahrscheinlich falsch, dazu gleich noch mehr):
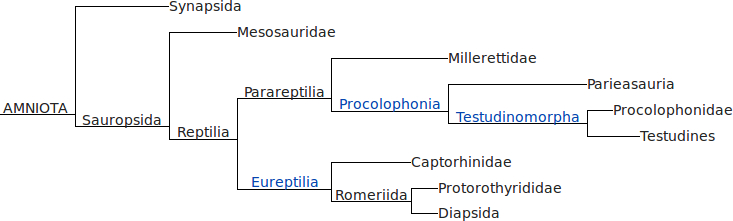
Oben sind die Synapsiden – die Säugetiere und ihre Verwandten und Vorfahren. Dann kommen die Mesosaurier – wasserlebende, sehr urtümliche “Ur-Reptilien”. Und daran schließen sich die beiden großen Gruppen der eigentlichen Reptilien an – die Parareptilien und die “echten” Reptilien (Eureptilia).
Zu den Parareptilien gehörten auch die Pareiasaurier, urtümliche und meist etwas plump aussehende Tiere. Einer davon war Bunostegos. Schaut man sich die Vorderbein-Knochen von Bunostegos aber genauer an, stellt man fest, dass er gar nicht so plump war, wie man dachte. Anders als bei den anderen Pareiasauriern liegen das Schulter- und das Ellbogengelenk nämlich so, dass der Oberarmknochen gerade unter dem Körper gehalten wurde. Bunostegos lief also nicht mit abgespreizten Beinen, sondern aufrecht. Bunostegos ist damit – nach bisherigem Wissen – das erste Landtier, das diese Gangart entwickelte, die später zum Erfolg von Dinosauriern und Säugetieren beitrug.
Auch ein anderes sehr urtümliches Reptil wartet mit Überraschungen auf: Captorhinus aguti sitzt ganz unten am Stammbaum der echten Reptilien. Untersucht man aber seine Knochen, erlebt man eine Überraschung: Man findet nämlich keine Wachstumslinien, wie sie typisch für viele Reptilien sind (und auch bei Säugetieren vorkommen). Captorhinus zeigt solche Linien aber nicht, was (nach Ausschluss anderer Möglichkeiten wie der, dass das untersuchte Fossil zu jung war) darauf hindeutet, dass die Tiere kontinuierlich und möglicherweise vergleichsweise schnell wuchsen.
Das Laufen auf zwei Beinen ist ja auch etwas, das man eher mit Dinosauriern und ihren unmittelbaren Vorfahren (ja, auch mit Menschen – pfffft, Säugetiere…) in Verbindung bringt. Aber auch hier haben die urtümlichen Parareptilien zuerst mit dieser Idee herumprobiert: Eudibamus ist ein Fossil aus Deutschland, das lange Hinterbeine besitzt, mit denen das Tier zumindest wenn es schnell unterwegs war, auf zwei Beinen laufen konnte. Computersimulationen zeigen, wie genau die Hinterbeine das anstellen – allerdings standen die Beine nicht senkrecht unter dem Körper wie bei Dinos (oder bei Bunstegos), sondern waren etwas abgespreizt. Ganz elegant war der Lauf von Eudibamus also wohl nicht.
Danach ging es um Schildkröten. Und hier gab es gleich zwei Untersuchungen, die sich mit Eunotosaurus beschäftigten, einem Reptil, das man standardmäßig bei den Parareptilien eingeordnet hat (auch Wikipedia tut das), das aber viele Merkmale mit Schildkröten gemeinsam hat und das man deswegen auch für einen Vorfahr der Schildkröten hält. (Heutzutage stützt man so etwas natürlich mit einer kladistischen Analyse.) Ein wichtiges Merkmal, das Schildkröten mit den Parareptilien verbindet, ist der “anapside” Schädel – ein Schädel, bei dem es hinter dem Auge keine weitere Schädelöffnung mehr gibt. Die meisten Reptilien dagegen haben solche Öffnungen und sind “diapsid”. Eunotosaurus hat nun – wie eine neue Untersuchung mittels CT-scan herausfand – ebenfalls eine solche Öffnung im Schädel – die hat man nur bisher nicht gesehen, weil sie durch einen Knochen, der eigentlich weiter hinten im Schädel liegt (Supratemporale) überdeckt wird. Die Knochen, die bei den anderen Diapsiden die Schädelöffnung bilden, haben auch beim Eunotosaurus eine solche Öffnung – sie ist eben nur von oben durch das Supratemporale zugedeckt. Das spricht dafür, dass Eunotosaurus (und damit auch die Schildkröten) doch zu den Diapsiden gehört (was molekulargenetische Untersuchungen auch nahelegen). Der vollkommen geschlossene Schildkrötenschädel entwickelte sich dann später.
Der zweite Vortrag drehte sich dann um die Entwicklung der Atmung in Schildkröten – wegen ihres starren Panzers können die ja nicht so einatmen, wie wir das tun (indem sie den Brustkorb weiten). Eunotosaurus hatte schon ziemlich breite Rippen und einen relativ starren Brustkorb und dürfte schon auf ähnliche Weise geatmet haben. Mehr über dieses Thema findet ihr hier.
Am Samstag morgen gab es dann noch einmal Dinos – bisher hatte es ja nur eine Session über Theropoden gegeben, die ja immer sehr im Rampenlicht setehn. (Das merkte auch einer der Vortragenden an, als er über die verlängerten Halswirbel des von ihm neu gefundenen antarktischen Fossils eines Ornithopoden (also eines Verwandten von Iguanodon) sprach – er sagte sinngemäß “Das Tier hat versucht, seine Halswirbel zu verlängern, damit es den Saurischiern ähnlicher wird, weil die ja viel populärer sind”.)
Zuerst ging es allgemein um die Gehirngröße von Dinos – generell gelten die ja als eher kleinhirnig, aber das zu quantifizieren ist nicht so einfach, denn je größer ein Tier wird, desto kleiner ist seine Hirnmasse im Verhältnis zur Körpergröße. Um hier also einen sinnvollen Vergleich zu haben, muss man sich überlegen, welche Hirngröße man bei einer bestimmten Körpergröße erwartet. Und als zweites kommt hinzu, dass es natürlich nicht einfach ist, die Hirngröße eines Dinos überhaupt abzuschätzen – das Gehirn muss in den Schädel passen, aber füllt es das Innere der Hirnschale voll aus wie bei Vögeln, oder eher nicht, wie bei Reptilien? Neue Ansätze, bei denen CT-scans verwendet werden, um die Hirnform genauer abzuschätzen, können hier vielleicht helfen.
Anschließend ging es um die Nasen von Ankylosauriern (den Panzerdinos der Kreidezeit). Die hatten sehr komplizierte und lange Luftwege in der Nase. Mit Hilfe von Computermodellen wurde simuliert, dass solche Luftwege sich gut dazu eignen, um die Luft beim Einatmen anzuwärmen (und beim Ausatmen abzukühlen) und auch, um Wasser zurückzuhalten. Was in meinen Augen fehlte, war die Gegensimulation: Wie viel schlechter wären die Verhältnisse gewesen, wenn die Wege für die Luft kürzer und weniger verschlungen gewesen wären (wie es ja auch bei anderen Dinos der Fall war)? Ohne eine solche Gegenprobe ist ja schwer zu sagen, ob die langen Wege wirklich aus diesem Grund entwickelt wurden oder ob sie nicht zu etwas anderem gut waren (zum Beispiel, um den Geruchssinn durch große Oberfläche zu verbessern?).
Dann gab es einen Vortrag, in dem Fossilien unter dem Elektronenmikroskop analysiert und eine überraschende Vielzahl von biologischen Strukturen gefunden wurden – rote Blutkörperchen, weiße Blutkörperchen, Kollagenfasern. Ziemlich überraschend – und ich bin nicht so ganz überzeugt. Da in dem Vortrag aber explizit gebeten wurde, die Ergebnisse nicht zu verbreiten, da sie kurz vor der Publikation stehen, schreibe ich hier lieber nichts, das nicht auch im Abstract steht.
Für mich als jemand, der sich für biologische Materialien interessiert, war der Vortrag über die Zähne von Horndinosauriern besonders interessant. Die hatten ja ein richtiges Scherengebiss, bei dem die Zähne im Ober- und Unterkiefer senkrecht aneinander vorbeischnitten und dabei die Pflanzen kleinschnipselten. Die meisten Reptilien müssen ja mit ziemlich simplen Zähnen auskommen – außen herum der hoch keramikhaltige Zahnschmelz, drinnen das knochenähnliche Dentin (oder Orthodentin), also Zahnbein, während Säugetiere zusätzlich auch noch Zement (der hauptsächlich aus Kollagen besteht) haben – hmmm, die vierte im Vortrag erwähnte Zahnkomponente will mir gerade nicht einfallen. Einige Dinos aber konnten das noch toppen – von Entenschnabeldinos kennt man 6 unterschiedliche Materialien im Zahn, bei den Horndinos sind es immerhin 5. (Dass Dinos hier mehr Komponenten haben als Säugetiere, war der “running gag” des Vortrags – Sprüche der Art “Hadrosaurs have more components – not that we are counting, but six is larger than four” gab es einige Male zur großen Freude des Auditoriums.) Die Horndinosaurier ordneten diese Materialien so an, dass an der jeweiligen Innenseite des Scherengebisses, da, wo die Zähne direkt aneinander abgleiten, die Zahnfläche selbst aus weicherem Material war, während außen herum härteres Material lag. Dadurch wurden die Zähne innen ein wenig stärker abgenutzt und eine scharfe Schneidkante blieb jeweils stehen.
Eher kritisch sehe ich den Vortrag, in dem es um die Wirbel der ganz großen Sauropoden (der Langhals-Langschwanz-Dinos) ging. Die sind typischerweise an einem Ende konvex, am anderen konkav (ein bisschen wie bei einem Kugelgelenk, wenn auch nicht so ausgeprägt). Um herauszufinden, ob diese Anordnung eher für hohe Stabilität oder für hohe Flexibilität sorgt, wurden Krokodilkadaver untersucht. Krokodile haben solche Wirbel nämlich auch im Rückgrat, aber nicht im Schwanz. Beim Manipulieren der Kadaver stellte sich nun heraus, dass der Schwanz von Krokodilen viel beweglicher ist als der Brustbereich – und daraus wurde geschlossen, dass diese Wirbelform eher für Stabilität sorgt. Nun ja – anzunehmen, dass alle Untersciede in der Beweglichkeit zwischen Schwanz und Körper nur an der Wirbelform liegen (und nicht etwa an Rippen, Knorpeln, dem umliegenden Krokodilpanzer etc.) ist schon etwas gewagt, um es vorsichtig auszudrücken.
Nachmittags ging es dann noch einmal um diverse Vierfüßer aus der Zeit vor den Dinos. Vielleicht lag es daran, dass ich nach so vielen Konferenztagen etwas abgestumpft war, aber die meisten dieser Vorträge fand ich etwas trocken. Es wurde versucht, urzeitliche Nahrungsketten zu rekonstruieren und zu sehen, wie empfindlich sie auf Störungen reagieren (das war ganz interessant, aber es gingen doch ziemlich viele Annahmen in die Überlegungen ein), diverse neue Fossilien wurden vorgestellt und einiges mehr.
Thematisch interessant (aber etwas sehr langweilig vorgetragen) war ein Vortrag über die Entwicklung des Ohrs bei frühen Säugetiervorfahren. Als die Fische an Land kamen, waren sie ja erst mal nicht für’s Hören in Luft ausgestattet – hören konnten sie allenfalls, wenn sie ihre Kiefer auf den Boden legten. Untersucht man die Orientierung des Hörknochens (der Steigbügel, wen ich mich nicht vertue), dann kann man daraus schließen, ob sich das Ohr eher zum Hören von Bodenschall oder von Luftschal eignet (und ein nettes Tierchen gab es auch, das vermutlich viel Zeit in seinen Bauten verbrachte und deswegen auch gut für das Hören von Schall ausgestattet war, der auf die Kopfoberseite übertragen wurde).
Und auch der letzte Vortrag war nett – hier ging es um die Entwicklung der Säugetierzähne (die haben aber nur maximal 4 Materialien – wollte ich nur erwähnen…). Bei Reptilien und den Säugetiervorfahren gab es ja viele Zahnwechsel – allerdings mit dem Nachteil, dass die Zähne dann natürlich nicht so perfekt ineinandergreifen, wie sie das bei Säugetieren tun. (Eine Ausnahme sind einige Dinos, wie oben ja schon angemerkt, eine andere ist zum Beispiel Pakasuchus.) Säugetiere haben ja typischerweise nur einen Zahnwechsel, und die Zähne wachsen eher langsam und spät, zumal sie ja auch direkt nach der Geburt eher stören würden (besonders die Mutter). Schaut man sich Zähne im Querschliff unter dem Mikroskop an, dann kann man in ihnen Bänder erkennen, die das tägliche Wachstum im Zahnschmelz wiedergeben (im Zahnbein gibt es solche Bänder auch, die wurden aber noch nicht angeguckt.) Und damit kann man dann herausbekommen, ob die Zähne schnell oder langsam wachsen. Wenig überraschend stellte sich heraus, dass bei den entfernteren Säugetiervorfahren die Zähne schnell wuchsen, während die ersten Fast-Säugetiere (untersucht wurde Oligokyphus) eher langsam wachsende Zähne hatten. Als nächstes wird man dann Zwischenformen angucken, um zu sehen, wann genau sich die Wachstumsgeschwindigkeit wie geändert hat.
Ja, und das war der letzte Vortrag bei der diesjährigen SVP-Tagung. Danach gab es noch eine Postersession und abends eine Preisverleihung – aber die Poster hatte ich mir schon mittags angeschaut und so bin ich dann wieder Richtung Heimat verschwunden, wo ich dann irgendwann am späten Abend erschöpft, aber auch ziemlich zufrieden, ankam.
SVP 2014 – ja, ich war dabei!





Kommentare (6)