Aber wie funktioniert Apoptose? Wie zerstören sich die Zellen selbst? Das ganze ist, wie so vieles in der molekularen Zellbiologie, sehr kompliziert und keineswegs vollständig erforscht. Ich bescheide mich daher damit, hier zwei Hauptarten vorzustellen ohne allzusehr ins Detail zu gehen: die extrinsische, von außen ausgelöste und die intrinsische, von der Zelle selbst ausgelöste Apoptose.
Die extrinsische Apoptose ist für den Körper die Möglichkeit, nicht mehr benötigte oder gar gefährliche (z.B. infizierte) Zellen zu entfernen. Im Immunsystem zum Beispiel gibt es cytotoxische T-Zellen, die mit einem Virus infizierte Zellen von außen erkennen können und dann Botenstoffe (=Liganden) abgeben, die an ein Empfängermolekül das alle Zellen auf ihren Oberflächen tragen (=Rezeptor) binden und damit in der Zelle eine Kaskade von Prozessen in Gang setzt, an deren Ende die Apoptose steht.
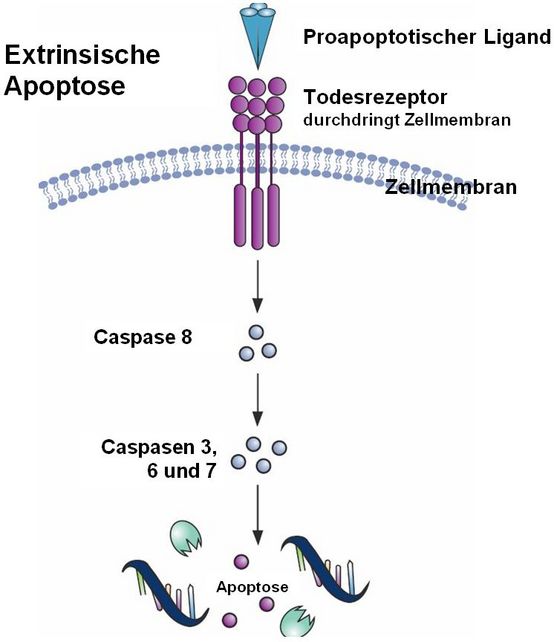
Im Bild sieht man, wie der Todesrezeptor einer Zelle ein Signal von außen erhält, es durch die Zellmembran nach innen weiterleitet und so die Apoptosekaskade initiiert. Ganz wichtig für die Ausführung der Apoptose sind dabei die sogenannten Caspasen, Enzyme, die gezielt andere Proteine an bestimmten Stellen zertrennen und damit zerstören können. Caspasen sind also gefährlich für die Zellen und müssen daher gut kontrolliert und gesichert sein: sie funktionieren erst, wenn sie „angeschaltet” werden.
Genau das geschieht durch die extrinische Apoptoseeinleitung. Hierbei wird immer die Caspase 8 aktiviert. Diese wiederum aktiviert dann die Caspasen 3, 6 und 7 – das sind die sogenannten Effektorcaspasen, die dann in der Zelle sozusagen die Abrissarbeiten übernehmen, indem sie das zelluläre Inventar darunter auch die DNA zerlegen.
Bei der intrinsischen Apoptose ist das ganze noch deutlich komplizierter.
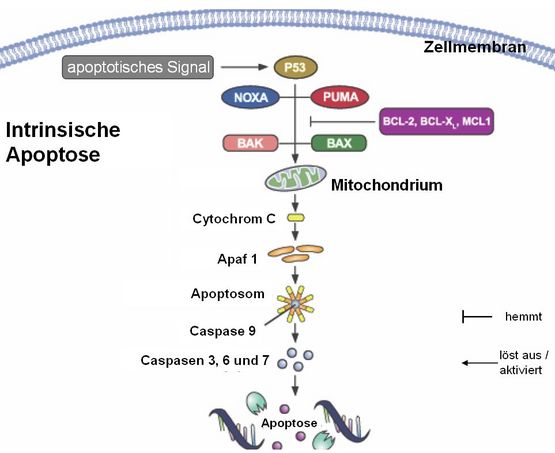
Hier löst ein zellinterner Stimulus die Apoptose aus. Dieser Stimulus kann vieles sein, z.B. ein unphysiologischer pH-Wert, ein Mangel an Glucose oder irreparabel beschädigte DNA (z.B. durch UV-Licht oder Chemikalien). Bestimmte Proteine, die die Beschädigung registrieren können, z.B. das „berühmte” p53 signalisieren dann: „hat keinen Zweck mehr. Abschalten!”. Ob letztlich tatsächlich die Apoptose eingeleitet wird, ist dann noch abhängig von einer Vielzahl von Einflussgrößen (, deren detaillierte Beschreibung den angestrebten Rahmen dieses Artikels pulverisieren würde). Im Bild sieht man eine Reihe von Proteinen, die diesen zellulären Signalweg modifizieren (Noxa, Puma, Bak, Bax) und einige andere, z.B. BCL-2, die die Apoptose hemmen können (was unter bestimmten Umständen ebenfalls sinnvoll sein kann). Erst wenn nach Integration aller zellulären pro- und antiapoptotischen Signale der proapoptotische Stimulus überwiegt (das ist nicht ganz unähnlich dem „Entscheidungsprozess” in Nervenzellen, darüber ob ein Aktionspotential ausgelöst wird oder nicht, indem Signale aus hemmenden und erregenden Nervenzellen zusammengefasst werden), wird das Mitochondrium zur Freisetzung von „Cytochrom c” veranlasst.
Danach gibt es dann kein Zurück mehr: zusammen mit dem Protein Apaf 1 bildet sich ein Komplex, das sogenannte „Apoptosom”, welches die Caspase 9 aktivieren kann. Die so scharf geschaltete Caspase 9 aktiviert ihrerseits die Effektorcaspasen, die wir oben schon kennengelernt haben und wir wissen ja jetzt, was diese anrichten: die Zelle stirbt.
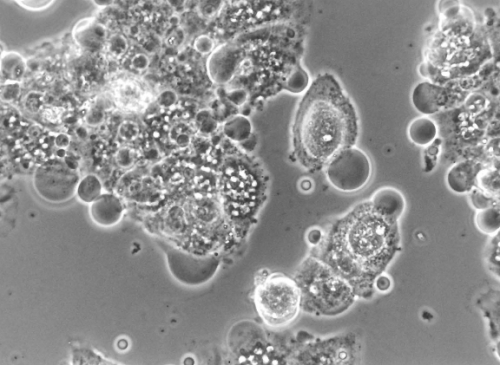
So wie auf diesem Bild sehen apoptotische Zelle aus. Sie schrumpfen und runden sich ab und man erkennt noch das „membrane blebbing” (sieht ein bißchen so aus, als würde die Zellmembran Blasen werfen), die Kondensation des Chromatins, die Fragmentierung des Zellkerns und einen Verlust der Adhäsion. Es bilden sich die typischen “apoptotischen Körperchen”, die sich Makrophagen und/oder benachbarten Zellen einverleiben (= phagozytieren). Im Inneren der Zelle, nicht mikroskopisch beobachtbar aber biochemisch nachweisbar, werden die DNA und intrazelluläre Proteine und Strukturen in Stücke zerlegt. Was also von der Zelle übrig bleibt ist, ist sehr überschaubar, recyclebar und – das ist wichtig – unschädlich für die Umgebung der Zelle.
Die sehr schematische Darstellung dieser beiden Wege der Apoptoseeinleitung sollte übrigens nicht dazu verleiten, sie sich als völlig isoliert voneinander vorstellen, denn es gibt durchaus Schnittstellen und Wechselwirkungen zwischen ihnen, die – wie so oft in der Biologie – alles noch viel komplizierter machen. Hinzu kommt, daß es mit der sogenannten „Nekroptose” ohnehin ein Phänomen gibt, das insofern eine Schnittmenge von Apoptose und Nekrose darstellt, als es Charakteristika von beiden aufweist. Darüber hinaus kennt man noch die Prozesse der Autophagie, bei der die Zelle Teile von sich selbst verdaut, und der Seneszenz, ein “Alterseffekt” bei Zellen, der u.a. verhindert, daß sie sich weiter teilen, die ebenfalls Mechanismen sind, die zur Optimierung der Resourcenverwaltung aber auch zum Schutz vor Infektionen und Entartung dienen und deren Abläufe und Regulation dicht in das zelluläre Kommunikationsnetz eingeflochten und auch von der Apoptosesteuerung nicht scharf zu trennen sind.
Wir stellen also fest, daß Funktionalität und “Konformität” der Zelle einer strengen und straffen Kontrolle sowohl von innen als auch von außen unterliegen und daß die reinigende, formende, schützende und nachhaltige Funktion der evolutiv stark konservierten programmierten Selbstzerstörung der Zellen untrennbar mit Gesundheit und Überleben des gesamten Organismus verbunden ist.
Wir werden später noch sehen, daß das, was unser Actionheld am Anfang dieses Beitrages tut, nämlich den Selbstzerstörungsmechanismus zu stoppen, in einer Zelle einen der Grundsteine der Krebsentstehung legen würde…
![]()

{kind=link}
Kommentare (3)