In der letzten Folge haben wir bei einem Ausflug ins Genom die kurzen DNA-Abschnitte short tandem repeats (STR) kennengelernt und ich habe gezeigt, wie und warum man sie für die Erstellung eines DNA-Profils benutzen kann.
In dieser Folge erkläre ich, wie man bis zu 16 STR-Systeme gleichzeitig mittels Multiplex-PCR vervielfältigen kann und wie der Cocktail aus verschiedenen DNA-Schnipseln durch Kapillarelektrophorese sortiert wird. (Ich empfehle dringend, sich vorher ein wenig über die PCR zu erkundigen.)
STR sind also unsere Zielsequenzen: die Kombination der Genotypen seiner STR-Systeme ist für jeden Menschen einzigartig (Ausnahme: eineiige Zwillinge). Aber wie stellt man fest, welchen Genotyp, also welche Allelkombinationen ein Mensch für seine verschiedenen STR-Systeme aufweist?
Zunächst einmal müssen wir genug Kopien der STR-Bereiche haben, um überhaupt damit arbeiten zu können. Im Falle einer sehr geringen Spurenmenge, z.B. einigen wenigen Hautzellen, die ein Täter an einem Messergriff hinterlassen hat, reicht das vorhandene Material für eine direkte Untersuchung der STR nicht annähernd aus und außerdem stört dabei auch der ganze Rest des normalen zellulären Genoms, das wir gar nicht anschauen wollen (und dürfen – s. StPO §81).
Eine kleine Analogie: ein Jahrmarktkünstler hat den ganzen Brockhaus (Genom) auf ein Reiskorn (Spurenträger) geschrieben und gibt uns das Korn. Wir sind aber nur interessiert an den 16 Einträgen über die deutschen Bundesländer (16 STR-Systeme), die über den ganzen Brockhaus verteilt sind. Da wir nur das eine Korn haben und nur einen Versuch, nehmen wir eine ganz starke Lupe und Papier und Stift zur Hand, vergrößern die Abschnitte die uns interessieren und schreiben sie jeweils auf ein Blatt Papier (PCR). Mit den 16 abgeschriebenen Seiten, die sich so leicht handhaben, ablesen und vervielfältigen lassen, können wir nun problemlos weiterarbeiten.
Die Abbildung zeigt die Verteilung der 13 STR-Systeme der amerikanischen CODIS über die einzelenen Chromosomen: überall im Genom finden sich STR-Systeme.
Um mit möglichst wenig DNA zurecht zu kommen und so auch materialarme Spuren bearbeiten zu können, wurden für die gleichzeitige Anreicherung von derzeit bis zu 16 STR-Systemen „Multiplex(=mehrfach)-PCR”-Verfahren entwickelt, die eine parallele, also simultan im selben Reaktionsgefäß stattfindende Vervielfältigung von 16 STR-Systemen gestattet. Für die Vervielfältigung eines bestimmten Bereichs des Genoms werden immer je zwei Primer (s. PCR) benötigt, für eine 16er Multiplex-PCR bedarf es daher 32 Primer, die genau für diesen Zweck designt und aufeinander abgestimmt werden müssen.
Und es kommt noch dicker: bei 16 STR-Systemen, deren beide Allele (eins auf jedem Chromsom eines Paars) je einen von etlichen möglichen Werten aufweisen können, ist die Wahrscheinlichkeit, daß Allele von unterschiedlichen STR-Systemen gleich groß sind, sehr hoch. Zum Beispiel:
Die Abbildung zeigt zwei DNA-Stücke, das eine gehört zu STR1, das andere aber zu STR2, die die gleiche Anzahl von Basen besitzen (s. DNA) und deshalb nicht mehr nur an ihrer Länge unterscheidbar wären.
Um solche PCR-Produkte, die von verschiedenen STR-Systemen stammen und dennoch die gleiche Länge haben, auseinanderhalten zu können, werden sie mit einem Fluoreszenzfarbstoff (Fluorophor) markiert und zwar immer so, daß zwei gleich lange Fragmente einen (von bis zu fünf) unterschiedlich farbigen Fluorophor tragen. Das funktioniert, indem die Fluorophore direkt an die Primer gebunden werden, die im Verlaufe der PCR ja in das entstehende Produkt eingebaut werden, so daß jedes einzelne Amplifikat eine von fünf Farben trägt. Da man vorher weiß, welche Allelwerte in welchen STR-Systemen auftreten können, kombiniert man STR-Systeme und Farben so, daß ausgeschlossen ist, daß zwei gleich lange Fragmente verschiedener STR-Systeme auch dieselbe Farbe tragen. So könnte eine solche Zuordnung aussehen:
Am Ende einer Multiplex-PCR liegt also immer ein Gemisch vor, in dem jedem Allel aller darin vervielfältigten STR-Systeme einem DNA-Fragment mit einer einmaligen Kombination von Länge und Farbe entspricht. Die Abbildung zeigt ein Beispiel:
Obwohl die Fragmente gleich lang sind, sind sie nun an ihrer Farbe zu unterscheiden.
Die Ermittlung der Größe eines Fragments schließlich gelingt durch einen Vergleich mit einem internen Größenstandard. Man kann sich das wie ein Lineal vorstellen, das man benutzt, um einen Faden unbekannter Länge zu messen, indem man den Faden daneben hält. Ein Größenstandard in unserem Fall ist eine künstlich hergestellte Mischung aus DNA-Fragmenten bekannter Größe, die natürlich alle, wie die vorvorige Abbildung zeigt, einen eigenen Farbstoff tragen (hier “lila”), der nicht für eines der getesteten STR-Syteme verwendet wird, damit DNA-Fragemente des Größenstandard und der zu untersuchenden Probe klar auseinandergehalten werden können.
Die stark vereinfachte Abbildung zeigt, wie man durch Vergleich mit den Fragmenten eines Größenstandard die Größe der beiden Allele des STR-Systems 1 bestimmen kann. Die “Zacken” repräsentieren hier jeweils ein DNA-Fragment: in diesem Beispiel hätten die Fragmente/Allele des STR-Systems 1 also die Größen 150 bp und 250 bp.
Eine solche Multiplex-PCR zu entwicklen, zu optimieren und als „Kit” marktreif zu machen, ist eine Kunst für sich. Es müssen geeignete Primer designt und aufeinander abgestimmt werden und zwar so, daß jede Einzel-PCR in etwa die gleiche Effizienz aufweist, die Kombinationen von Längen und Farbstoffen müssen berechnet, ein geeigneter Größenstandard muß hergestellt und eine optimale „Chemie” (ein Gemisch aus Salzen, Stabilisatoren und Lösemitteln, die die Grundlage der chemischen Reaktion bildet) muß zusammengestellt werden. Angesichts der höchsten Ansprüche, die solche Kits erfüllen und angesichts der peniblen und umfangreichen Validierung, die sie dafür durchlaufen müssen, ist das insgesamt ein langwieriges, aufwendiges und teures Unterfangen, das sich die Hersteller, die solche Kits für forensische STR-Multiplex-PCRs anbieten, auch sehr gut bezahlen lassen.
Nach der Multiplex-PCR liegen jedoch erst einmal alle Fragmente wild gemischt in einem einzigen Reaktionsgefäß vor, von einem geordneten DNA-Profil sind wir so noch weit entfernt. STR-Allele unterscheiden sich, wie erwähnt, in der Anzahl ihrer Wiederholungseinheiten. Dadurch sind sie unterschiedlich lang und genau dieser Längenunterschied soll gemessen werden. Und zwar auf das einzelne Nukleotid genau, da ja auch die sogenannten „Punktallele” (s. hier), die nur ein Nukleotid länger oder kürzer als ein anderes Allel sein können, voneinander unterschieden werden müssen. Um diese Fragmente so exakt der Größe nach zu ordnen, ihre Größe zu messen und ihre Farbe erkennen zu können, müssen wir nun eine Kapillarelektrophorese durchführen.
Das Prinzip der Elektrophorese ist irgendwann einen eigenen Basics-Artikel wert, daher nur so viel: DNA-Fragmente sind in wässriger Lösung negativ geladen und bewegen sich in einem elektrischen Feld daher immer zum positiv geladenen Pol (Anode). Da bei der DNA die Linienladungsdichte konstant und die Anzahl der Ladungen proportional zur Länge ist, bestimmt die Länge eines DNA-Fragments, wie rasch ein solches zur Anode wandert: je länger ein DNA-Fragment ist, desto langsamer wandert es. Gibt man ein Gemisch von unterschiedlich langen DNA-Fragmenten in eine gleichmäßig aufgebaute Matrix (wie ein Netz) und legt daran eine Spannung an, so beginnt ein „Wettrennen” der DNA-Fragmente, die sich durch die Matrix drängen und sich exakt gemäß ihrer Länge hintereinander „einreihen”.
Genau dieses Prinzip nutzt man bei der Kapillarelektrophorese: die Matrix, hier ein bestimmtes, hochwertiges Polymer, ist in einer haardünnen Kapillare untergebracht, die in das Reaktionsgefäß eingetaucht wird. Durch Elektroden wird ein starkes elektrisches Feld erzeugt, so daß die Fragmente in die Kapillare eintreten und durch das Polymer Richtung Anode wandern können.
Quelle: https://www.pharmchem.tu-bs.de/
An einer Stelle der Kapillare befindet sich ein „Fenster”, eine durchsichtige Öffnung, durch die ein Laserstrahl geschossen wird. An diesem Fenster wandern die Fragmente, schön der Größenreihe nach, vorbei. Das Laserlicht trifft sie, regt die Fluorphore, die an die Fragmente gebunden sind, zum Leuchten in ihrer jeweiligen Farbe an und dieses Leuchten wird von einem gegenüberliegenden Detektor registriert. Somit werden die beiden „Datenpunkte” Länge und Farbe erhoben und einem bestimmten Fragment zugeordnet.
Eine Software erstellt aus diesen Rohdaten und unter Einbeziehung der bekannten Werte für den Größenstandard ein Elektropherogramm.
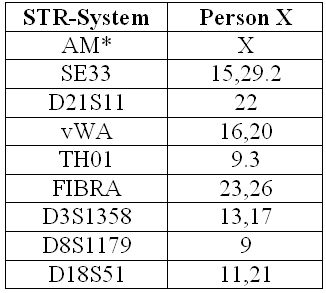
Die Abbildung zeigt ein Elektropherogramm mit 15 autosomalen STR-Systemen und dem geschlechtsspezifischen Amelogenin-System (AM). In den grünen Balken steht die Bezeichnung des jeweiligen STR-Systems. Die „Zacken” nennt man „Peaks” und sie repräsentieren jeweils ein DNA-Fragment bestimmter Größe. Jedem STR-System sind ein oder zwei Zacken zugeordnet, je nachdem, welche Allelkombination ein Mensch im betrachteten STR-System besitzt.
Die Beschriftung der Peaks übernimmt die Software, denn sie „weiß”, welche Allele in welchem STR-System vorkommen können und wie lang diese Allele sind. Sie integriert also die Informationen „Länge”, „Farbe”, „Zuordnung einer Farbe zu möglichen STR-Systemen” und „Allelidentitäten und -größen in STR-System” zu einer eindeutigen Bezeichnung eines Peaks als Allel X, der Größe Y aus STR-System Z.
Bei dem Menschen, dessen Elektropherogramm die Abbildung zeigt, handelt es sich z.B. um einen Mann (X und Y im Amelogenin-System), der im System D10 die Allele 12 und 15, im System vWA die Allele 14 und 16 aufweist usf.
Aus einem solchen Elektropherogramm läßt sich endlich sehr einfach ein DNA-Profil, wie es hier schon gezeigt wurde, ableiten.
In der nächsten Folge unserer beliebten Serie machen wir einen Ausflug in die Populationsgenetik und ich erkläre, warum ein DNA-Profil aus 16 STR-Systemen tatsächlich mit hoher Wahrscheinlichkeit einzigartig auf der Welt ist.
![]()

{kind=link}
Kommentare (18)