Man lerne in der Schule zwar dem Vernehmen nach nichts über das (Alltags)Leben, dafür aber werde dort ausdrücklich nicht unerwähnt gelassen, daß Mitochondrien die Kraftwerke der Zellen seien.
Diese Bezeichnung, die vermutlich wirklich die allermeisten schon einmal gehört haben, stellt auf eine der wichtigsten Funktionen dieser Organellen ab, die man in (fast) allen eukaryotischen Zellen, bei Pflanzen, Pilzen und Tieren, findet: sie wandeln die “Rohenergie” aus der Nahrung in ein chemisches Energieäquivalent um, das überall und in allen Zellen des Körpers benötigt wird, um Prozesse, die nicht von selbst ablaufen würden (endergon), mit der notwendigen Energie zu versorgen und dieses Äquivalent, diese “Energiewährung” heißt Adenosintriphosphat (ja, das Zeug, aus dem u.a. auch die DNA besteht), kurz “ATP”.
Das ist zwar sowohl sehr interessant als auch sehr wichtig und die entsprechenden biochemischen Details der Atmungskette konnte wahrscheinlich so ziemlich jeder Biologe und Biochemiker irgendwann während seiner Ausbildung auch nachts um vier geweckt abspulen. Dennoch ist dieser Prozess für die Forensische Genetik eher nebenrangig und darum soll es hier auch nicht gehen. Uns interessiert viel mehr die DNA der Mitochondrien, die mtDNA (s. Abbildung).

Schematische Darstellung eines Mitochondriums
Man erkennt die Doppelmembran und im Inneren eigene, ringförmige DNA-Moleküle
DNA? Wieso hat denn so ein Organell, so ein Zellbestandteil überhaupt eigene DNA? Das erklärt sich durch die Herkunft der Mitochondrien: gemäß der bestbelegten Theorie waren die Vorgänger der Mitochondrien selbstständige Urbakterien, die natürlich auch ein eigenes Genom hatten und die vor hunderten Millionen bis Milliarden von Jahren von anderen Urzellen phagozytiert wurden, wodurch eine Endosymbiose entstand, in der beide Organismen von den Fähigkeiten des jeweils anderen profitierten. Und auch heute noch zeigen Mitochondrien deutliche Anzeichen ihrer bakteriellen Herkunft, darunter ihre Doppelmembran und eben das Vorhandensein ihrer eigenen DNA, die sogar eine leicht andere genetische Kodierung als die nukleäre DNA (nDNA) verwendet. Inzwischen sind die Mitochondrien längst keine selbstständigen Organismen mehr und viele ihrer Gene sind im Lauf der Evolution in die nDNA versetzt worden, doch noch immer enthält ihre ringförmige DNA eine ganze Reihe lebenswichtiger Gene die dichtgedrängt und zusammen mit einigen nicht-kodierenden Sequenzen insgesamt 16.569 Basen lang sind (s. Abbildung). Unter diesen Genen ist auch Cytochrom b, das ich schon an anderer Stelle erwähnte, und welches forensisch interessant ist, wenn, wie in jenem Artikel erläutert, im Rahmen einer Ermittlung die Spezies, die eine bestimmte Spur hinterlassen hat, bestimmt werden soll. Dies kommt zwar in der normalen Routinearbeit nur sehr selten vor, ist jedoch extrem relevant für die Wildlife Forensik, die ich schon einmal erwähnt habe und worüber ich sicher noch einen eigenen Artikel schreiben werde.
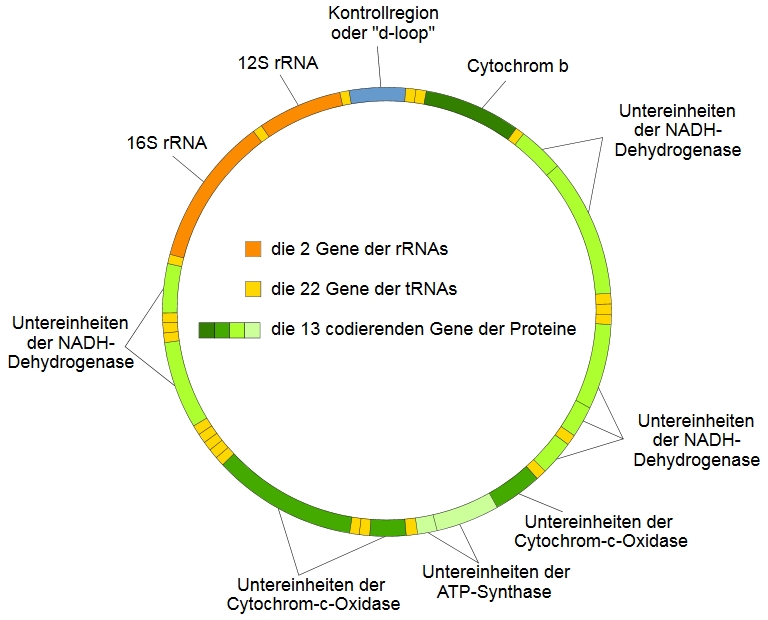
[a]
In der Forensik nutzt man diese Erkenntnis beispielsweise, wenn die DNA-Spuren an einem Tatort so beschädigt oder so minimal sind, daß das Standard-Verfahren für DNA-Profiling, das auf der Darstellung von STR-Systemen beruht, nicht mehr funktioniert. Der Vorteil der mtDNA gegenüber nDNA besteht darin, daß sie durch ihre geringe Größe und geschlossene Ringform viel stabiler ist, als die viel größere nDNA, so daß sie überdauern kann, selbst wenn die nDNA z.B. durch Umwelteinflüsse längst jenseits der Analysierbarkeit zerstört wurde. Außerdem gibt es in jeder Zelle hunderte bis tausend Mitochondrien, die jeweils mehre Kopien ihrer mtDNA enthalten. So findet sich in biologischem (= zellhaltigem) Spurenmaterial also immer tausende Male mehr mtDNA als nDNA, weshalb man, selbst wenn in einer Spur keine nDNA mehr nachweisbar ist, oft noch mtDNA darin finden und untersuchen kann.
Warum man dann nicht gleich immer die mtDNA für forensische Analysen untersucht? Das hat zunächst rein praktische Gründe: eine Sequenzierung ist aufwendiger, schwieriger und teurer als eine STR-Analyse. Die mtDNA hat zudem entscheidende Nachteile, die ihre forensische Brauchbarkeit deutlich einschränkt: sie ist ein Haplotyp, d.h. sie wird, so wie sie ist, sozusagen en bloc, weitervererbt, ohne daß Rekombination stattfindet und somit die für die enorme Diskriminierungsfähigkeit der STR-DNA-Profile erforderliche kombinatorische Vielfalt entstehen kann. Außerdem wird sie stets und ausschließlich über die mütterliche Linie vererbt: die Kinder einer Mutter erhalten immer das komplette mitochondriale Genom der Mutter, die mtDNA des Vaters wird nie vererbt*. Das bedeutet, daß alle Personen, die in mütterlicher Linie verwandt sind, theoretisch die gleiche mtDNA-Sequenz haben, was eine forensische Ermittlung erheblich erschweren kann, wenn für eine Tat beispielsweise auch der Bruder und/oder die Schwester eines Tatverdächtigen in Frage kommt, man aber nur das mtDNA-Profil als Vergleichsbasis hat. Hier ein Beispiel aus einem hypothetischen Mordfall:
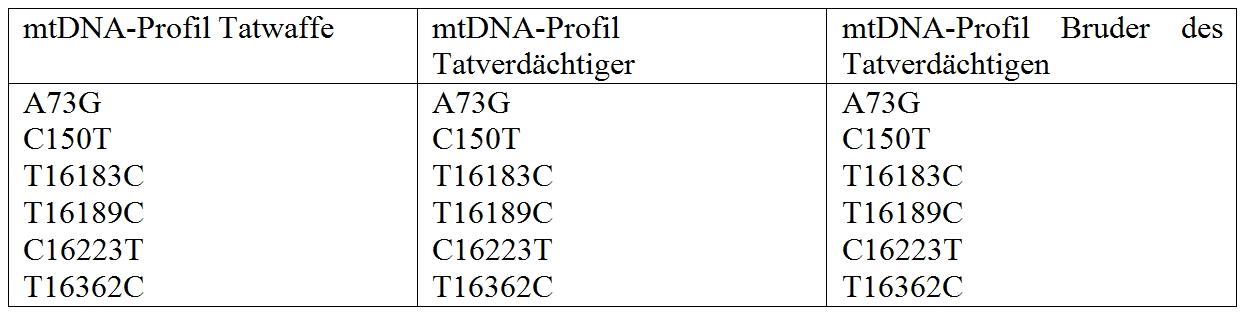
Die mtDNA an der Mordwaffe ist identisch mit der des Tatverdächtigen: sie zeigt dieselben Abweichungen von der Referenzsequenz, z.B. ein “G” statt einem “A” an der Position 73. Der Bruder des Tatverdächtigen, der auch als Täter in Frage kommt, hat jedoch das gleiche mtDNA-Profil
Aber selbst wenn nur eine einzige Person in Frage kommt und ihr mtDNA-Profil mit demjenigen aus der Spur an einer Mordwaffe übereinstimmt, besteht immer noch die Möglichkeit, daß die Übereinstimmung zufällig ist, weil manche mtDNA-Haplotypen sogar relativ häufig in der Bevölkerung bzw. bestimmten Subpopulationen vorkommen. Daher ist die mtDNA in forensisch-genetischen Ermittlungen immer eher ein letzter Rettungsanker, kann aber immerhin sicher den Ausschluß einer verdächtigen Person von der Täterschaft begründen, wenn deren mtDNA-Profil nämlich von dem eines entscheidenden Asservates eindeutig abweicht.
Aber auch in der Abstammungsbegutachtung, einer weiteren Routinedisziplin der forensischen Genetik, kann die mtDNA nützlich sein, indem man mit ihr eine mütterliche Linie verfolgen kann. Das kann hilfreich sein, wenn man es mit komplexeren als den üblichen Fragestellungen etwa nach der Vaterschaft eines Mannes zu einem Kind zu tun hat. Ein Beispiel für eine solche wären zwei Menschen, die wissen wollen, ob sie Halbgeschwister sind und von derselben Mutter abstammen oder unverwandt sind. Mit den standardmäßig verwendeten STR-Systemen gelangt man in solchen Fällen häufig nicht zu einer befriedigenden Antwort. Bezieht man die mtDNA ein, erhält man bei nicht übereinstimmenden Profilen einen sicheren Ausschluss einer gemeinsamen Mutter und bei übereinstimmenden Profilen einen weiteren mehr oder weniger starken Hinweis auf eine Halbgeschwisterschaft. Wie stark der Hinweis ist, hängt von der Seltenheit des gemeinsamen mtDNA-Haplotypen ab, die man unter Zurateziehung öffentlicher Datenbanken wie EMPOP ermitteln oder aber zumindest abschätzen kann.
Ein weiteres Beispiel für eine komplexe Abstammungsbegutachtung wäre folgendes: Eine Großmutter “X” möchte wissen, ob ihre Enkelin “Z” von ihrer Tochter “A”, die sie mit einem anderen Mann als ihren Sohn “Y” gezeugt hat und die demnach eine Halbschwester von “Y” ist, abstammt oder von “B”, die mit ihr, der Großmutter zwar unverwandt, jedoch eine Halbschwester von “Y” ist und mit diesem einen gemeinsamen Vater hat. Weder A noch B sind jedoch verfügbar für einen Test und da die Beteiligten zu einer sehr kleinen, nach außen quasi isolierten ethnischen Gruppe gehören, die Verhältnisse in dieser Familie sehr “unübersichtlich” sind und laut Y durchaus auch B als Mutter von Z in Frage kommt, ist das auf STR-Systemen beruhende Ergebnis der Begutachtung, in die nur X, Y und Z einbezogen wurden, nicht zufriedenstellend.

blaue Personen haben einen gemeinsamen mtDNA-Haplotypen; gestrichelte Linien deuten eine mögliche aber nicht bestätigte Verwandtschaft an
Hier kann die mtDNA helfen: wenn Z wirklich über A von X abstammt, muß Z eine identische mtDNA wie X haben. Dies war nicht der Fall, also konnte A als Mutter ausgeschlossen werden. Aber auch für Fragestellungen, die über konkrete Abstammungsverhältnisse weit hinaus gehen und die Genealogie über zahlreiche Generationen betreffen, spielt die mtDNA eine wichtige Rolle. So kann mtDNA-Haplotypen so weit zurückverfolgen, daß man sogar von einer mitochondrialen Eva spricht.
Die mtDNA ist also in der forensischen Genetik eher eine Helferin in speziellen und schwierigen Fällen und gar nicht so viele Labore, die von der DAkkS nach der DIN 17025 für die Durchführung von gerichtlich anerkannten STR-Analysen akkreditiert sind, haben auch eine Akkreditierung für mtDNA-Analysen (wir hier haben sie übrigens) oder nehmen am entsprechenden Teil der GEDNAP-Ringversuche teil.
Es gibt dennoch forensische Labore, die sich regelrecht auf mtDNA in Anwendung und Forschung spezialisiert haben, zu nennen ist hier vor allem die Forensische Genetik der Gerichtsmedizin Innsbruck, unter Leitung meines Kollegen W. Parson. Von dort stammen nicht nur die EMPOP-Datenbank und die Anwendung quasi-medianer Netzwerke zur Qualitätskontrolle von mtDNA-Haplotypen, sondern auch Arbeiten zur NGS-basierten Analyse der mtDNA [1] sowie zahlreiche Arbeiten mit das rein forensische Terrain verlassenden Aspekten, wie z.B. zoologischen, phylogenetischen, populationsgenetischen und genealogischen Fragestellungen. Derselben Gruppe gelang damals auch und ebenfalls mittels mtDNA-Analyse sowohl die Entlarvung des Schädels, der 180 Jahre lang in Schillers Sarg gelegen hatte, als Fälschung [2], als auch die Identifikation der 2007 in Jekaterinenburg gefundenen Gebeine als Überreste der bislang vermissten Zarenkinder Alexei und seiner Schwester Maria Romanowa, nachdem bereits 1996, allerdings von einer anderen Gruppe, ebenfalls via mtDNA, die Authentizität der Gebeine des Zaren Nikolaus II. nachgewiesen worden war [3].
Übrigens: auch in meiner Gruppe hat gerade ein Bachelor-Student der naturwissenschaftlichen Forensik ein Projekt zur Analyse von mtDNA unter ganz bestimmten forensischen Gesichtspunkten fertig gestellt, über das ich hier noch nichts aber bestimmt schon bald mehr erzählen kann. Die mtDNA wird uns indes wohl, auch wenn einige ihre forensische Relevanz durch Techniken wie NGS und Marker wie SNPs bedroht sehen, noch ein Weilchen erhalten bleiben.
____
*Exkurs: Es ist noch gar nicht so lange bekannt, wie es kommt, daß nur die mütterliche mtDNA in den Nachkommen eines Elternpaars vorhanden ist. Erst 2011 zeigten Sato&Sato und Rawi et al. [4,5], daß die väterlichen Mitochondrien aus einer befruchteten Eizelle durch Autophagie, einem Abbauprozess, der für die Zelle möglicherweise schädliche Strukturen entfernt, zerstört werden. Diese neuen Erkenntnisse lösen daher die vorherigen Erklärungsversuche ab, denen zufolge die schiere Überzahl mütterlicher Mitochondrien die väterlichen einfach bis zur Unauffindbarkeit verdünnen. Diese Befunde erklären übrigens nur, daß und wie väterliche Mitochondrien aus den Zygoten beseitigt werden. Warum das passiert, also, zu welchem Zweck, liegt nach wie vor im Dunkeln. Möglicherweise ist mtDNA-Heteroplasmie, also das gleichzeitige Vorliegen mehrerer verschiedener mtDNAs schädlich für den Embryo. Oder schaden bestimmte Faktoren aus den väterlichen Mitochondrien (also nicht deren DNA) dem Embryo? Vielleicht werden auch väterliche Mitochondrien (irrtümlich) von der weiblichen Eizelle als Pathogene/fremde Eindringlinge erkannt und deshalb angegriffen? Die Erkenntnisse von Sato und Rawi werden jedenfalls dazu beitragen, dieses Geheimnis zu lüften, was von großer Bedeutung für die menschliche Entwicklungsbiologie und das Verständnis mitochondrialer Erkrankungen sein wird.
______
Referenzen:
[1] Parson, W., Strobl, C., Huber, G., Zimmermann, B., Gomes, S. M., Souto, L., … & Irwin, J. (2013). Evaluation of next generation mtGenome sequencing using the Ion Torrent Personal Genome Machine (PGM). Forensic Science International: Genetics, 7(5), 543-549.
[2] „Die vertauschten Köpfe“, Der Spiegel 19/2008
[3] Ivanov, P. L., Wadhams, M. J., Roby, R. K., Holland, M. M., Weedn, V. W., & Parsons, T. J. (1996). Mitochondrial DNA sequence heteroplasmy in the Grand Duke of Russia Georgij Romanov establishes the authenticity of the remains of Tsar Nicholas II. Nature genetics, 12(4), 417-420.
[4] Sato, M., & Sato, K. (2011). Degradation of paternal mitochondria by fertilization-triggered autophagy in C. elegans embryos. Science, 334(6059), 1141-1144.
[5] Al Rawi, Sara, et al. “Postfertilization autophagy of sperm organelles prevents paternal mitochondrial DNA transmission.” Science 334.6059 (2011): 1144-1147.
__
Bildquelle:
[a] layout by jhc – https://commons.wikimedia.org
![]()

{kind=link}
Kommentare (9)