Man lerne in der Schule zwar dem Vernehmen nach nichts über das (Alltags)Leben, dafür aber werde dort ausdrücklich nicht unerwähnt gelassen, daß Mitochondrien die Kraftwerke der Zellen seien.
Diese Bezeichnung, die vermutlich wirklich die allermeisten schon einmal gehört haben, stellt auf eine der wichtigsten Funktionen dieser Organellen ab, die man in (fast) allen eukaryotischen Zellen, bei Pflanzen, Pilzen und Tieren, findet: sie wandeln die “Rohenergie” aus der Nahrung in ein chemisches Energieäquivalent um, das überall und in allen Zellen des Körpers benötigt wird, um Prozesse, die nicht von selbst ablaufen würden (endergon), mit der notwendigen Energie zu versorgen und dieses Äquivalent, diese “Energiewährung” heißt Adenosintriphosphat (ja, das Zeug, aus dem u.a. auch die DNA besteht), kurz “ATP”.
Das ist zwar sowohl sehr interessant als auch sehr wichtig und die entsprechenden biochemischen Details der Atmungskette konnte wahrscheinlich so ziemlich jeder Biologe und Biochemiker irgendwann während seiner Ausbildung auch nachts um vier geweckt abspulen. Dennoch ist dieser Prozess für die Forensische Genetik eher nebenrangig und darum soll es hier auch nicht gehen. Uns interessiert viel mehr die DNA der Mitochondrien, die mtDNA (s. Abbildung).

Schematische Darstellung eines Mitochondriums
Man erkennt die Doppelmembran und im Inneren eigene, ringförmige DNA-Moleküle
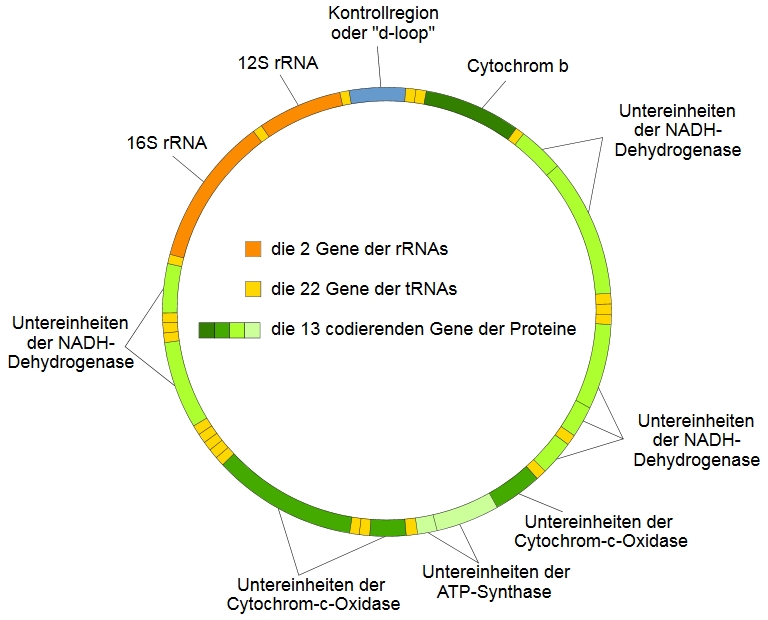
DNA? Wieso hat denn so ein Organell, so ein Zellbestandteil überhaupt eigene DNA? Das erklärt sich durch die Herkunft der Mitochondrien: gemäß der bestbelegten Theorie waren die Vorgänger der Mitochondrien selbstständige Urbakterien, die natürlich auch ein eigenes Genom hatten und die vor hunderten Millionen bis Milliarden von Jahren von anderen Urzellen phagozytiert wurden, wodurch eine Endosymbiose entstand, in der beide Organismen von den Fähigkeiten des jeweils anderen profitierten. Und auch heute noch zeigen Mitochondrien deutliche Anzeichen ihrer bakteriellen Herkunft, darunter ihre Doppelmembran und eben das Vorhandensein ihrer eigenen DNA, die sogar eine leicht andere genetische Kodierung als die nukleäre DNA (nDNA) verwendet. Inzwischen sind die Mitochondrien längst keine selbstständigen Organismen mehr und viele ihrer Gene sind im Lauf der Evolution in die nDNA versetzt worden, doch noch immer enthält ihre ringförmige DNA eine ganze Reihe lebenswichtiger Gene die dichtgedrängt und zusammen mit einigen nicht-kodierenden Sequenzen insgesamt 16.569 Basen lang sind (s. Abbildung). Unter diesen Genen ist auch Cytochrom b, das ich schon an anderer Stelle erwähnte, und welches forensisch interessant ist, wenn, wie in jenem Artikel erläutert, im Rahmen einer Ermittlung die Spezies, die eine bestimmte Spur hinterlassen hat, bestimmt werden soll. Dies kommt zwar in der normalen Routinearbeit nur sehr selten vor, ist jedoch extrem relevant für die Wildlife Forensik, die ich schon einmal erwähnt habe und worüber ich sicher noch einen eigenen Artikel schreiben werde.
[a]
In der Forensik nutzt man diese Erkenntnis beispielsweise, wenn die DNA-Spuren an einem Tatort so beschädigt oder so minimal sind, daß das Standard-Verfahren für DNA-Profiling, das auf der Darstellung von STR-Systemen beruht, nicht mehr funktioniert. Der Vorteil der mtDNA gegenüber nDNA besteht darin, daß sie durch ihre geringe Größe und geschlossene Ringform viel stabiler ist, als die viel größere nDNA, so daß sie überdauern kann, selbst wenn die nDNA z.B. durch Umwelteinflüsse längst jenseits der Analysierbarkeit zerstört wurde. Außerdem gibt es in jeder Zelle hunderte bis tausend Mitochondrien, die jeweils mehre Kopien ihrer mtDNA enthalten. So findet sich in biologischem (= zellhaltigem) Spurenmaterial also immer tausende Male mehr mtDNA als nDNA, weshalb man, selbst wenn in einer Spur keine nDNA mehr nachweisbar ist, oft noch mtDNA darin finden und untersuchen kann.
![]()

{kind=link}
Kommentare (9)